Project 18. Mechanistic insights into co-transcriptional mRNA packaging complexes.
Nuclear mRNA export is an essential step in eukaryotic gene expression, ensuring that mature transcripts are delivered from the nucleus to the cytoplasm for translation1. mRNAs are transcribed as pre-mRNAs and are heavily processed, including capping, splicing and polyadenylation, prior to their nuclear export. These processing steps lead to the deposition of protein marks on the mRNA (such as the cap binding complex, the exon junction complex or polyA-binding protein), which in turn aid the packaging of the emerging ribonucleoprotein complex (mRNP) through RNA–protein and protein–protein interactions1. Packaging of the nascent mRNA is essential to prevent entanglement of the RNA with DNA, which would otherwise lead to the formation of R-loops at the site of transcription, resulting in genome instability. Here, a key player is the TREX complex, consisting of the DExD-box ATPase UAP56 and the THO complex2, which aids co-transcriptional mRNP packaging and helps establish a ‘maturation vessel’ for the forming RNP3.
Transcription by Polymerase II is pervasive. In contrast, only a small fraction of transcripts yields mature, fully processed mRNAs for nuclear export4. However, RNP packaging is essential for all transcripts to prevent the formation of harmful R-loops at the site of transcription. RNA packaging presumably relies on the engagement of mRNA processing marks and mRNA export components, such as the TREX complex5. This creates a series of challenges: How is the co-transcriptional packaging of all PolII transcripts ensured? How are only fully matured mRNAs are exported? How are mRNAs discriminated against ‘faulty mRNAs’ and transcription byproducts, which are targeted for degradation? To shed light on these questions we here aim at studying mRNA packaging complexes, and in particular address the following questions:
- How is the RNA packaging machinery recruited to all PolII transcripts?
- How are maturing mRNAs saved from being mis-interpreted as transcription byproducts and funneled into mRNA export instead of degradation?
- How does an imbalance between nuclear RNA degradation and export affect R-loops at the site of transcription?
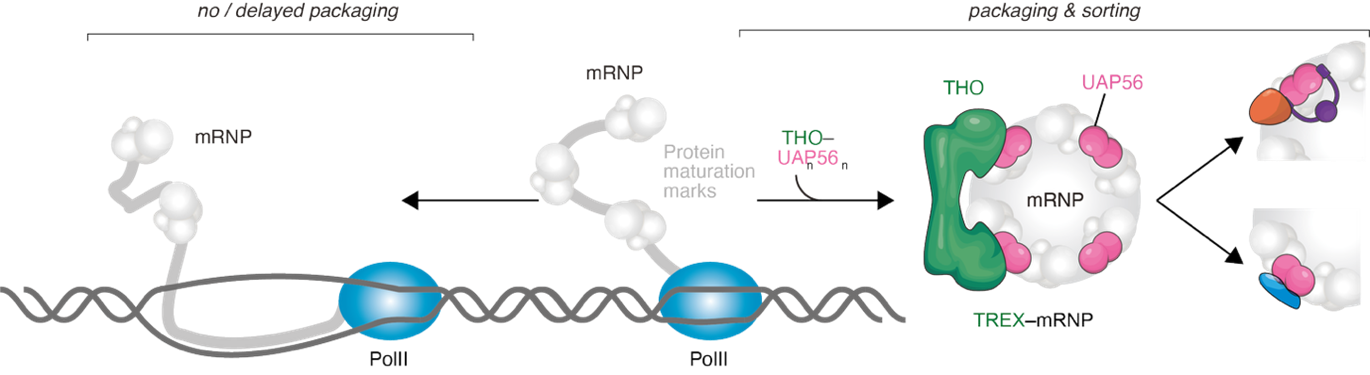
Using a combination of biochemical reconstitution, biophysical characterization and structural biology (including AlphaFold predictions and cryo-Electron Microscopy), we will investigate the formation of novel mRNP packaging complexes. We recently identified various novel protein-protein interactions between mRNP components. We hypothesize that these help (1) shield the mRNP from both degradation and export, directly promoting an increased window for mRNP packaging and (2) constitute a ‘fast-track’ complex towards mRNP formation and nuclear export. Applying cellular assays (endogenous CRISPR-tagging, reporter systems, RNA-FISH), proteomics, transcriptomics and high-resolution microscopy we will further dissect the co-transcriptional recruitment of the packaging and export machinery to nascent RNAs.
Taken together, by combining cryo-EM, biochemical reconstitution, and in vivo assays, we will elucidate new mechanisms contributing to co-transcriptional mRNP packaging and sorting. Our work will reveal how coordinated mRNA maturation counteracts R-loops at the site of transcription and acts as an essential guardian of genome stability.